NEONATOLOGY ON THE WEB
Historical Review and Recent Advances
in Neonatal and Perinatal Medicine
Edited by George F. Smith, MD and Dharmapuri Vidyasagar, MD
Published by Mead Johnson Nutritional Division, 1980
Not Copyrighted By Publisher
Chapter 14
Researches in Perinatal Circulation
John Lind, Professor Emeritus
INTRODUCTION
Goeffery Dawes, the famous British physiologist has presented the
unborn in the following way (1969):[1] "The human
fetus has been likened to a spaceman; passive, insulated and
preserved from stimuli, which is only half the truth. On the
contrary, one could think of the mammalian embryo as a hitch-hiker
with a large pack on his back getting into a rather small car; he is
a friendly fellow who chatters away all the time and is prepared to
do some driving if given half a chance -- he takes you off your route
and tells when and where he would like to get out." In the same way,
the fetal circulatory system, different as it is from the child's or
the adult's, cannot be dismissed with the common term "immature;" it
is a system well adapted to the circumstances, and at no stage is
there any evidence of any inadequate circulatory competence.
THE FETAL CIRCULATION
The first recorded mention of the fetal cardiovascular system is
attributed to Galen who, in the Second Century A.D., described what
were later to become known as the foramen ovale and its valve, as
well as the ductus arteriosus, and gave some account of their
post-natal closure.[2]
Galen writes: "Nature is neither lazy nor devoid of foresight.
Having given the matter thought, she knows in advance that the lung
of the fetus, a lung still contained in the uterus and in the process
of formation and spared continual motion, does not require the same
arrangements of a perfected lung endowed with motion. She has
therefore anastomosed the pulmonary artery with the aorta, and the
left and right atria. . . ."
Falloppio (1561) initiated the special use of the word "placenta"
(literally a flat cake or pancake),[3] and three
years later, in 1564, a posthumous publication by Vesalius contained
the first account of the ductus venosus.[4] In
1626, Spigel, also in a work published after his death, pointed out
the absence of any direct communication between the umbilical vessels
of the fetus and the uterine vessels of the mother. He also noted the
fact that, in the fetus, the two ventricles are of approximately
equal thickness, whereas in the adult the left ventricle
predominates.[5]
In 1628, William Harvey introduced his concept of the circulation
of the blood and included in his treatise the first account of the
fetal circulation.[6] The publication of his work
heralded the beginning of the use of a dynamic total concept of the
cardiovascular system as opposed to the anatomical descriptions of
separate individual structures which had been available up to then.
Harvey realized that in the fetus the two ventricles work in parallel
instead of in series as in the adult. He appears, however, to have
denied the existence of any pulmonary circulation at all in the
fetus. In the year 1652, Olof Rudbeck[7]
demonstrated for Queen Christina of Sweden and her court his
discovery of the lymph vessels.
In the seventeenth century, several other capital discoveries in
human physiology were made. In 1660, Robert
Boyle[8] demonstrated that part of the air is
essential to life. The following year, Malpighi produced the first
descriptions of capillaries and of terminal airspace' in the
lungs.[9] Six years later, Hooke proved that
respiration depends on adequate supply of fresh air to the
lungs.[10] In the same year, Walter Neddham called
the placenta the "uterine lung." Lower (1669) verified experimentally
Hooke's postulate and also showed that venous blood owes its dark
colour to "loss of air."[11]
In the following centuries, the course of the fetal circulation
was a subject of speculation and controversy. The main problems
discussed were the distribution of the superior and inferior caval
flow through the fetal heart and whether or not there was a
significant pulmonary blood-flow. No notable progress in our
understanding was made until the modern era of physiological
investigation was started in the twentieth century of Pohlman (1909).
He injected starch into different fetal vessels and studied the
distribution of the granules.[12]
Hugget (1927) found that the oxygen content of carotid arterial
blood was greater than that of umbilical arterial
blood.[13] In 1928 Kellog[14]
initiated experiments in which the fetus was delivered by caesarean
section with the fetal and maternal circulation retained and the
fetal respiration prevented.
Forsmann, the heroic genius, carried out the first
heart-catheterization and angiocardiography on himself standing
before a mirror during flouroscopy.[15] The year
was 1929. Nobody dared to assist him. These two methods meant an
explosive progress in the study of the circulation.
Few men of science have contributed so much to the advancement of
physiology in the first half of this century as Sir Joseph Barcroft.
One of his many talents was the ability to attract younger
colleagues. In 1938, Barcroft and Barron united their experience of
fetal physiology with Barclay, Franklin and Prichard's experience in
radiography. The combined Cambridge and Oxford teams obtained the
first direct records of the course of the circulation in the fetal
lamb.[14]
In the concluding portion of their book, The Foetal
Circulation, the authors discussed the possibility of (for what
was then the future) direct studies of the human fetal
circulation.[16] They expressed the opinion that
the best hope for the achievement of this lay in the use of
angiocardiographic technique suitably modified and refined. Such
studies have been carried out in connection with legal abortions and
have resulted in a confirmation of the findings in fetal lambs (Lind
and Wegelius 1954).[17] They also pointed out the
reciprocal relationship between abnormal respiratory adaptation and
cardiovascular disturbances in the neonatal period.
However, angiocardiography is not a quantitative technique. It
was, therefore, highly desirable to obtain such information. In 1939,
Dawes and his associates, working at the Nuffield Institute for
Medical Research in Oxford, presented data on the normal distribution
of the blood-flow, derived from the oxygen saturation of blood
withdrawn simultaneously from various vessels, in the mature fetal
lamb exteriorized by caesarean section.[18] Among
the many new data presented, it should be mentioned that the total
pulmonary blood-flow averages only about 10% of the combined
ventricular output. The quantities of blood flowing through the
foramen ovale, ductus arteriosus and aortic isthmus are large,
between 35 and 45% of the combined ventricular output. About 55%
streams through the placenta, which is characterized by low vascular
resistance.
The changes in circulation at birth were first described by Dawes
and coworkers: the sudden drop in pulmonary resistance with an
increase in pulmonary blood-flow; a reversal in the direction of
blood-flow through the ductus arteriosus and the closure of the
foramen ovale (Born et al., 1954), a result of the greatly increased
blood-flow through the lungs to the left
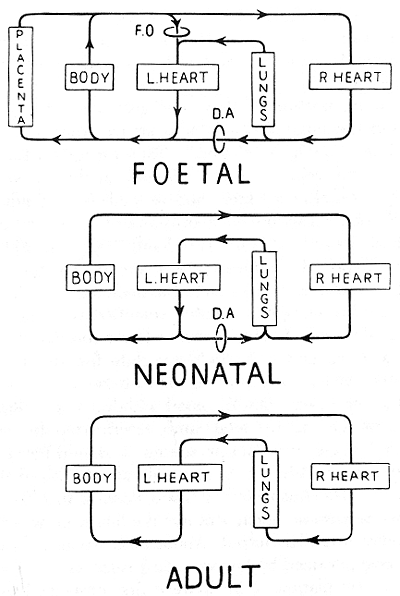
atrium[19] (Fig. 1).
The studies of the fetal circulation in lambs and the changes at
birth were pursued by Rudolph and coworkers using more physiological
methods (1967). They used indwelling catheters and examined the fetal
and newborn lambs in utero and in resting state without use,
of anesthesia.[20] They could follow the increase
in combined cardiac output as well as the changes in actual
blood-flow to various organs during gestation.[21]
Rudolph presented fundamental data on the post-natal changes in
cardiac output and its distribution. He found that the right
ventricle in the lamb fetus in late gestation ejects about two thirds
of the combined ventricular output or about 300 ml/kg/min. With the
elimination of the placental circulation and the decrease in
pulmonary vascular resistance, the right ventricle ejects all its
blood into the pulmonary circulation. When the ductus arteriosus
closes, this means about 200 ml/kg/min is ejected, an amount similar
to that which has passed through the placenta in utero. The left
ventricle which prenatally ejects about 150 ml/kg/min, or one-third
of the combined ventricular output, increases its output after birth
about 25% to 200 ml/kg/min.[22]
THE IMMEDIATE CHANGES IN THE CIRCULATION AT BIRTH
Our concept regarding the pre-natal circulation in humans is based
mainly on animal studies. There seems to be little reason to question
the validity of the basic pattern of fetal circulation derived from
these experiments, and a limited number of observations in the human
fetus corroborate these assumptions. The knowledge about the
immediate circulatory adaptation to extrauterine life is also largely
built on data from prenatal and postnatal animal studies but to a
great extent is also based on studies of the human circulation in the
postnatal period.
THE ESTABLISHMENT OF PULMONARY VENTILATION
The proper airfilling of the lungs in the immediate neonatal
period has two major functions:
1. It will allow for the exchange of 02 and
C02, a function carried out in fetal life by the
placenta.
2. The dynamic pressure-flow alterations produced by the
expansion of the lungs provide the mechanism around which the
newborn's circulatory patterns are initiated, maintained, and
varied in the transitional period.
These two functions are so closely related that they should not be
thought of as two separate processes. The circulatory adjustment
represents a total integrated cardio-pulmonary response, upon the
smooth and successful accomplishment of which the fate of the newborn
depends. That they are considered separately, is done only for the
object of clarity.
INITIATION OF BREATHING AND DYNAMICS OF LUNG LIQUID
The fetal lungs are solid organs secreting fluid into the
bronchial tree. They extract oxygen from the blood instead of
contributing to oxygenation.
It was Jost (1934) from Paris who first provided evidence of the
formation of fluid in the fetal lung of the rabbit. When the trachea
was ligated, the lungs became distended over a period of
time.[23]
Forrest Adams, pediatrician at U.C.L.A., and his team (1963) made
extensive animal studies on the production and composition of the
lung liquid and found that it differed chemically from amniotic
fluid.[24]
Aeration of the lungs is thus not inflation of collapsed empty
organs but rather a replacement of intra-alveolar fluid by air. It
has been shown by X-ray that the human lungs occupy about the same
intrathoracic volume before and after the first breaths (Lind et al.,
1966).[25]
Karlberg et al. (1962)[26] demonstrated that
during vaginal delivery, the thorax is subjected to pressures up to
95 cm of H2O and some fluid is expressed from the lungs.
There is then an elastic recoil of the thorax which results in
aeration of at least the upper airways.
Active inspiration of 30-70 ml is achieved by the first
inspiration and most of this air remains to form part of the residual
volume of the lungs. This volume increases with subsequent breaths
and the functional residual air is 17 ml/kg body weight at 10
minutes, about twice that 20 minutes later, and shows no further
increase during the first week (Karlberg et al.,
1962).[26]
At birth, a rapid reversal of the direction of lung-liquid flow is
essential for smooth transition from placental to pulmonary
gas-exchange. Recent studies have shown that removal of liquid from
the lungs begins before birth (Lawson et al.,
1978;[27] Bland et al.,
1979)[28] and that the increase in cathecholamine
production during labor stimulates the absorption of lung liquid.
Intravenous infusion of epinephrine into late term fetal lambs causes
reabsorption of liquid from potential airspaces (Walters and Olver
1978).[29]
Substantial volume of fluid remains in the lungs even after the
first breaths and has to be removed. How this happens has been
studied in animal experiments by Leonard Strang and his colleagues at
University College, London[30] but it is still a
matter of discussion. When breathing starts, a transpulmonary
pressure-gradient develops which inflates the lungs and displaces
residual liquid from the terminal respiratory airspaces into the
distensible perivascular tissue spaces surrounding pulmonary blood
vessels distant from sites of respiratory gas exchange (Bland et al.,
1979).[31] The concentration of protein in the
lung tissue drops, thereby increasing the transvascular protein
gradient. Strang found a considerable increase in lymph flow,
following birth draining of the lung tissue. Bland and his group,
working at the Cardiovascular Research Institute in San Francisco
have, over the last years, performed a series of studies on the
perinatal lung fluid dynamics. In one study (1980), they measured the
excess liquid in the lungs of unanesthesized late term fetal lambs
before and during delivery. The volume late in the labor was only 5-7
ml/kg of body weight compared to 20 ml/kg before labor. After birth
the pulmonary lymph flow increased and the magnitude of drainage this
way was found to be 15-20% of the total lung liquid. Thus, most of it
is absorbed directly by the drastically increased microcirculation of
blood, which expands the effective vascular surface area for fluid
exchange in the lungs.[32]
THE ESTABLISHMENT OF THE POST-NATAL PULMONARY CIRCULATION
The high resistance of the fetal pulmonary vascular bed is
believed to be localized in the precapillary muscular arterioles
which develop a thick muscular coat during the latter part of
gestation (Civin and Edwards, 1951).[33] This
muscle mass permits them to function as sphincters controlling the
volume flow through the lungs. It regresses by about 40% at two weeks
of age (Naeye 1961).[34]
Cook and associates (1963)[35] and Cassin and
coworkers (1964)[18] stressed the importance of
low PO2 in maintaining pulmonary vasoconstriction in the
fetus, and the immediate increase in pulmonary blood flow associated
with ventilation of the lungs with gas containing oxygen.
Rudolph and Yuan (1966) studied the pulmonary vascular responses
to variation in blood oxygen tension and H+ ion concentration levels
in newborn calves.[22] They found a curvilinear
inverse relationship between pulmonary vascular resistance and
arterial Pot. The vasoconstrictive response to hypoxia was markedly
enhanced by lowering arterial pH (Fig. 2).
A reduction of pulmonary blood flow is of little consequence in
fetal life where oxygenation is carried out in the placenta. In the
neonatal period, however, the development of acidosis and hypoxia
from any cause can produce pulmonary vasoconstriction which can have
serious consequences. In the immediate neonatal period, a fetal
pattern of circulation with right-to-left shunting through the ductus
arteriosus and foramen ovale will be reestablished, increasing the
hypoxia and the pulmonary vascular resistance, thus creating a
vicious circle. The relationship between pulmonary vascular
resistance, PO2 and pH, was elucidated by Rudolph and
Yuan, which has been of fundamental importance for clinical practice
in neonatology.
Other factors also tend to reduce the vascular resistance and
increase in pulmonary blood flow after birth. Thus, the production of
a liquid-gas interface within the minor airspaces creates a surface
tension which can contribute to maintenance of lung vessel patency.
It has also been demonstrated that the mechanical effect or
respiration movement increases the pulmonary blood
flow.[18]
Among the various pharmacologically active pulmonary vasodilators
in the newborn period, a-blocking agents, Beta2-agonists and
prostaglandin 12 (prostacyclin) are worth mentioning (Lock et al.,
1979).[36]
THE CLOSURE OF THE DUCTUS ARTERIOSUS
During fetal life, the ductus arteriosus permits the right
ventricle to develop pari passu with the left side of the
heart, in spite of the very reduced lung perfusion. It is
questionable if the left-to-right shunt which starts with the abrupt
reduction in pulmonary vascular resistance is necessary or even
meaningful for the postnatal pulmonary circulation. The shunt can be
looked at as an additional help to establish an adequate perfusion of
the lungs and also as a safety-valve for sudden increase in pulmonary
vascular resistance, which otherwise could jeopardize the function of
the right ventricle. It must, however, be stressed that the existence
of a wide communication between the systemic and the pulmonary
circulation carries a risk of volume overloading the left heart from
hyperfusion of the lungs, in view of the fact that the left ventricle
in late fetal life is used to handle only one-third of the combined
ventricular output. An effective synchronization of the drop in
pulmonary vascular resistance and the reduction in caliber of the
duct seems a prerequisite for prevention of left ventricular failure.
Fig. 3 summarizes the resistance in the systemic and pulmonary
circulation and the size of the shunt through the ductus. The fact
that hardly any shunt can be detected 20 hours after birth, in spite
of a resistance ratio between systemic and pulmonary circulation on
the order of 2:1, denotes that the ductus is functionally closed
(Wallgren 1977).[37] Numerous studies have
demonstrated that the duct constricts in response to a variety of
stimuli but most researchers agree that the duct constricts at birth
primarily in response to high PO2, which was first
demonstrated by Kennedy and Clark (1941) at Vanderbilt
University.[38,39]
The original demonstration by Coceani and Olley
(1973)[40] that E-type prostaglandins are potent
relaxants of the ductus arteriosus, confirmed by subsequent animal
studies by others, in vitro (Starling and Elliott
1974)[41] and in vivo (Sharpe and Larsson
1975),[42] suggested their use in those cases of
congenital heart diseases was entirely dependent on persistence of
the ductus arteriosus for the maintenance of pulmonary blood flow. On
the other hand, indomethacin, a blocker of prostaglandin synthesis in
tissues, produces intense and persistent contraction of the ductus
arteriosus in vivo (Sharpe et al., 1974)[43] (Fig.
4). Indomethacin has, since then, been widely used clinically and has
proven an effective treatment when used on correct indications.
It is important to point out that the efficiency of ductal closure
is related to the maturity of the newborn and persistent patency is a
common finding in the premature infant, often complicating the
transition to extrauterine life.
CLOSURE OF FORAMEN OVALE
The foramen ovale is the main route for relatively direct passage
of blood from the placenta to the brain and normal development of the
left heart depends on patency of the foramen.
The closure of the foramen ovale is a consequence of the
hydrostatic condition following the increased lung perfusion. Because
of the initially small differences in the systemic and pulmonary
vascular resistance, and since the central circulatory pattern of the
newborn infant is labile-relatively small changes in pulmonary
vascular resistance will provoke relapses into fetal circulatory
pattern with ductal and/or atrial right to left shunting and arterial
desaturation.
Although functional closure is a rapid process (Fig. 5), which may
even occur with the first vigorous cry, anatomical closure is a slow
process which only exceptionally is complete at three months and
usually takes a year or more. Probe patency is seen in more than 50%
of children up to five years of age and in about 20% of adults.
PERINATAL SYMPATHO-ADRENAL ACTIVITY
The cardiorespiratory adaptation at birth demonstrates the great
capacity of the neonatal circulation to cope with all the strains
involved in the reorganization of the circulatory system postpartum.
Such a capacity suggests the existence of an effective system of
integrated baroreceptor and chemoreceptor reflexes (peripheral and
central).
It seems fair to state that the birth process constitutes a major
challenge to mechanisms controlling circulatory hemostasis in the
newborn. One function of the sympatho-adrenal system is to sustain
homeostasis in the newborn. Another function of the sympatho-adrenal
system is to sustain homeostasis during stress, and it is not
surprising that a strikingly increased activity of this system has
been recorded during birth. Lagercrantz et al., (1980) have recently
reviewed the sympatho-adrenal activity in the fetus during delivery
and birth.[44]
In the fetus and the preterm infants the sympathetic nervous
system might not be fully developed (Friedman 1973)45 but
the adrenal medulla is relatively voluminous (Comline et al.,
1966).[46] The fetus is also provided with
paraganglia containing a considerable amount of catecholamines.
Studies on previable human fetuses have demonstrated that hypoxia
causes a depletion of these paraganglia prior to that of adrenal
medulla (Hervonen et al., 1972).[47]
Adrenaline and particularly the noradrenaline concentrations are
high at birth after normal vaginal delivery, reaching levels ten
times higher than in the resting adult and also considerably higher
than in the mother during labor (Lagercrantz and Bistoletti
1977).[48] In asphyctic infants the values are
extremely high, probably because hypoxia and acidosis trigger their
release, as has been proven the case in laboratory fetuses (Jones et
al., 1975)[49] (Fig. 6). The squeezing of the
infant, particularly the head, in the birth canal seems to stimulate
their release, since very high values were seen in vacuum extracted
infants even in those without asphyxia. Furthermore, the
catecholamine concentrations seen after selective caesarean sections
are considerably lower than after uncomplicated vaginal deliveries,
probably due to lack of mechanical stress (Fig. 7).
The concentration of catecholamines in peripheral venous blood
remains fairly high during the first three hours after birth. Twelve
to 24 hours postnatally, the concentration decreases to normal
resting values in adults (Eliot et al., 1978).[50]
Catecholamines are certainly an asset to the newborn. They help to
redistribute the blood preferentially to the most vital organs-the
placenta, the heart and the brain (Dawes
1968).[51] They improve cardiac performance
(Downing et al., 1969)[52] and mobilize glucose
(Jones and Ritchie 1978).[49] They stimulate
absorption of the lung liquid and enhance the release of surfactant
(Lawson et al., 1978),[27] just to mention a few
of the vital contributions to neonatal cardiopulmonary function.
MYOCARDIAL STRUCTURE AND FUNCTION
Structural and functional studies of the fetal and neonatal lamb's
myocardium (Friedman 1972,[45] McPherson et al.,
1976)[53] demonstrated an inherent limitation of
contractility during this age period. A higher resting tension was
found in the fetal and neonatal lamb myocardium when compared with
that of the sheep. There were fewer contractile elements but more
nuclear material and water content in the myocardium of the fetus and
newborn. Rapid postnatal growth of the left ventricle occurs which
provides more "reserve" sarcomeres to handle the increasing blood
volume and end-diastolic pressure. This was demonstrated both
structurally and functionally in lambs (Riemenschneider et al.,
1978)[54] thus providing the basis for the
age-related improvement of the contractile response of the neonatal
myocardium to stretch according to Starling's law.
In the human newborn, Arcilla et al. (1966)[55]
have observed the absence of a post-extrasystolic potentiating effect
in ventricular contraction which contrasted with findings in older
children. This suggests a less well developed myocardial
contractility in the newborn. Based on findings of a higher
preejection period/left ventricular ejection time ratio, Hedvall
(1975)[56] concluded that left ventricular
function in the newborn infant is impaired.
An interesting finding is the increase in preejection period/left
ventricular ejection time ratio from birth to 4-to-6 hours of age in
neonates (Lundell and Wallgren 1981).[57] This
might be explained by the diminishing concentration of catecholamines
in the blood of the newborn.
Another possible explanation may be the relative decline in energy
substrate for the heart. The energy source for the myocardium is
conceivably influenced by the dramatic biochemical alterations during
and after birth. The blood glucose level in fullterm infants
decreases during the first 2 to 6 hours postnatally (Cornblath and
Schwartz, 1966)[58] when the infants are usually
fasting. This may depress myocardial function and contribute to the
increase in preejection period/left ventricular ejection time ratio
in infants at this age.
The amount of myocardial glycogen is normally high throughout
gestation (30-40 mg/g in the cardiac ventricles in human) (Fig.8) and
these stores increase at term (Shelley, 1961).[59]
In the presence of severe postnatal hypoxia, survival depends to a
certain extent on anaerobic energy release (Stafford and Weatherall,
1960).[60]
THE PLACENTAL TRANSFUSION
One circulatory parameter that has attracted a great deal of
interest in its physiological implication on the adaptive capacity of
the neonate is placental transfusion. When cord clamping is delayed
for 3-5 min. ("late-clamped") and blood volume (BV) determined 8-15
minutes after birth, there is an increment of about 33% compared with
infants whose cords were clamped within 10 sec. of birth
("early-clamped"), or a 25% increment in BV when measured at
1/2-hour-old (Oh et al., 1966).[61] Based on red
cell volume, a more satisfactory indicator of the magnitude of
placental transfusion, a 40-50% blood volume increase was estimated
in late-clamped infants (Usher et al., 1963).[62]
The late-clamped infant adjusts to a larger blood volume by
hemoconcentration, rapid plasma extravasation and increased urine
output during the first hours of life (Oh et al.,
1966).[63] Considerable differences in the
circulatory and respiratory function, between infants with small and
large placental transfusion, have been noted during the first days of
life.
The sudden augmentation in BV in late-clamped infants was observed
to be reflected even in the more distensible venous part of the
circulatory system. During the first hour of life, the atrial
pressures of the late-clamped infants are higher. Thereafter, the
difference between the two groups is not significant. The atrial
pressures of late-clamped infants drop to about half by the 2nd hour
or later, whereas that of the early-clamped infants remains unchanged
at comparable ages (Arcilla et al., 1966).[55] A
blood volume related pulmonary hypertension exists in the
late-clamped infants and a further hypertensive effect may be
elicited by the prevailing lower Pa02 and higher
PaC02 in these infants (Oh et al.,
1966).[61]
There is also a difference in systemic blood pressure between
early- and late-clamped infants (Oh et al.,
1966).[64] The latter have a relatively high
systolic pressure at 5 min. of life followed by a rapid decline
during the first 6 hours. In early-clamped infants, the systolic
pressure starts at a much lower level to be followed by a slight rise
during the first 15 min. Both groups show a low systemic blood flow
during the 1st day which increases by the 2nd or 3rd day, accompanied
by a decline in systemic vascular resistance.
Left-to-right shunts and, to a lesser degree, right-to-left shunts
persist longer postnatally in the late-clamped infants, suggesting
that closure of the fetal channels may occur later (Arcilla et al.,
1967).[65]
Unlike the early-clamped infants, the late-clamped ones are not
always entirely asymptomatic and expiratory grunting is not uncommon
(Yao et al., 1971)[66] during the first hours
after birth.
It has further been found that the preejection period is prolonged
in late-clamped infants suggesting that a sizable
placenta-transfusion affects adversely the left ventricular
performance of the neonate (Yao and Lind
1799).[67]
Concluding excuse: Ars longa -- vita brevis. This
review presents only relatively few studies of the overwhelmingly
rich literature on the perinatal circulation.
|

|
Figure 1. Dawes (1954) classic illustration of the
postnatal changes in the circulation.
|
|

|
Figure 2. The effects of changes in Pot, in pH alone and
in both combined, on the pulmonary vascular resistance as
derived from studies in newborn calves (Rudolph and Yuan
1966).
|
|

|
Figure 3. The shunt through the ductus arteriosus and the
relationship between systemic and pulmonary resistance
(Wallgren 1977).
|
|

|
Figure 4. Inner ductal diameter (X ± S.D.) is shown
immediately after delivery and up to 120 min. afterwards in
control pups and those exposed in utero to indomethacin by
dosing the dam either 18 or 12 h prior to delivery. Small
numbers denote the number of determinations at each point
(Sharpe, Thalme and Larsson 1974).
|
|

|
Figure 5. Oxygen saturation of the peripheral blood in
relation to birth of the infant (Wallgren 1977).
|
|

|
Figure 6. Catecholamine concentrations and pH values in
fetal scalp blood samples during labor and in umbilical
artery at birth (0 min.): (a) Six normal deliveries with
fetal pH <7.25; (b) six complicated deliveries with pH
<7.25.
|
|

|
Figure 7. Catecholamine concentrations in umbilical
arterial blood at birth, after uncomplicated vaginal
delivery, elective and emergency caesarean sections.
(Lagercrantz and Bistoletti unpublished data)
|
|

|
Figure 8. Cardiac carbohydrates and survival
during.anoxia (Stafford and Weatherall 1960).
|
REFERENCES
1. Dawes G. S.: Foetal autonomy. In Wolstenholme G. E. W. and
O'Connor M. (eds): A Ciba Foundation Symposium. London: J. and
A. Churchill.Ltd., p. 316, 1969.
2. Galen: Opera Omnia IV:243. In Dalton J. C.
(translator): Doctrines of the Circulation. Philadelphia:
Lea's Sons & Co, 1884, p. 68. Quoted from Rashkine W. J.:
A brief historical perspective. Pediatr. Cardiol. 1:62,
1979/80.
3. Falloppio B.: Observationes anatomicae. Ulmum M.
A., Venetiis, 1561.
4. Vesalius A. (1514-64): De Humani Corporis Fabrica
Libri Septem. Ex Off. Ioannis Oporim, Basileae 1543.
5. Spigel A.: Adriani Spigelii Bruxellensis equitis D.
Marci, olim in patavino Gymnasio Anatomiae et Chirurgiae Profess.
Primarij, De Humani Corporis Fabrica Libri Decem. Tabulis XCIIX aeri
incisis elegantissimis, nee ante hac visis exornati, screnissimo
Ioanni Carnelio Venetiarum Duci Dicati. Opus posthumum. Daniel
Bucretius Vratislaviensis Philos.
et Medic. D. Jussu Authoris in lucem profert.
Venetiis MDCXXVII.
6. Harvey W.: Movement of the heart and blood in
animals. Franklin K. J. (translator). Oxford: Blackwell
Scientific Publishers, 1957.
7. Rudbeck 0.: Nova exercitatio anatomica exhibens
ductus hepaticus aquosus et vasa glandulorum serosa. Uppsala,
1653.
8. Boyle R.: A defense of the doctrine touching the spring
and weight of the air. In The Work of the Honourable Robert Boyle.
London: p. 157-160, 1772.
9. Malpighi M.: De pulmonibus observations anatomicae. Two
letters (the second placed and dated Bologna, 1661) to
Borelli. Young J. (translator). Proc. R. Soc. Med. 23: Sect.
Hist. Med. 1-11, 1929.
10. Hooke R.: Lectures de potentia restitutiva, or of spring. In
Early Science in Oxford, Vol. 8, p. 333. Oxford: R. T.
Gunther, 1931.
11. Lower R.: Tractatus de Corde. Londini: J. Allestry,
1669.
12. Pohlman A.: The course of the blood flow to the heart
of the fetal mammal with a note on the reptilian and amphibian
circulations. Anat. Rec. 3:75, 1909.
13. Hugget A. St G.: Foetal blood-gas tensions and gas
transfusion through the placenta of the goat. J. Physiol.
(Lond) 62:673, 1927.
14. Barclay A. E., Barcroft J., Barron D. H. et al.: A
radiographic demonstration of the circulation through the heart in
the adult and in the foetus, and the identification of the ductus
arteriosus. Br. J. Radiol. 12:505, 1939.
15. Forssman W. T. O.: Die Sondierung des rechten Herzens.
Klin. Wochenschr. 8:2085, 1929.
16. Barclay A. R., Franklin K. J., Pritchard M. M.: The
foetal circulation and cardiovascular system, and the changes that
they undergo at birth. Oxford: Blackwell, 1944.
17. Lind J., Wegelius C.: Human fetal circulation: Changes
in the cardiovascular system at birth and disturbances in the
postnatal closure of the foramen ovale and ductus arteriosus. In
Cold Spring Harbor Symposia on Quantitative Biology, Vol. XIX, p.
109. New York, 1954.
18. Cassin S., Dawes G. S., Ross B. B.: Pulmonary blood
flow and vascular resistance in immature foetal lambs. J. Physiol.
(Lond) 171:80, 1964.
19. Born G. V. R., Dawes G. S., Mott J. C., et al.: Changes
in the heart and lungs at birth. In Cold Spring Harbor Symposia on
Quantitative Biology, Vol. XIX. New York 1954.
20. Rudolph A. M., Heymann M. A.: The circulation of the
fetus in utero. Methods for studying distribution of blood flow,
cardiac output and organ blood flow. Circ. Res. 21:163, 1967.
21. Rudolph A. M., Heymann M. A.: Circulatory changes with
growth in the fetal lamb. Circ. Res. 26:298, 1970.
22. Rudolph A. M., Yuan S.: Response of the pulmonary
vasculature to hypoxia and H± ion concentration changes. J.
Clin. Invest. 45:399, 1966.
23. Jost A.: Hormonal factors in the development of the
fetus. In Cold Spring Harbor Symposia on Quantitative Biology,
Vol. XIX. New York, 1954.
24. Adams F. H., Moss A. J., Fagan L.: The tracheal fluid
in the fetal lamb. Biol. Neonate. 5:151, 1963.
25. Lind J., Tahti E., Hirvensalo M.: Roentgenologic
studies of the size of the lungs of the newborn baby before and after
aeration. Ann. Paediatr. Fenn. 12:20, 1966.
26. Karlberg P., Adams F. H., Geubelle F., et al.:
Alteration of the infant's thorax during vaginal delivery.
Physiological studies. Acta. Obstet. Gynecol. Scand. 41:223,
1962.
27. Lawson E. E., Brown E. R., Torday J. S., et al.: The
effect of epinephrine on tracheal fluid flow and surfactant efflux in
fetal sheep. Am. Rev. Respir. Dis. 118:1023, 1978.
28. Bland R. D., Bressack M. A., McMillan D. D.: Labor
decreases the lung water content of newborn rabbits. Am. J.
Obstet. Gynecol. 135:364, 1979.
29. Walters D. V., Olver R. E.: The role of catecholamines
in lung liquid absorption at birth. Pediatr. Res. 12:239,
1978.
30. Strang L. B.: Uptake of liquid from the lungs at the
start of breathing. In de Renck A. V. S. and Porter R. (eds):
Development of the Lung. London: J & A Churchill Ltd,
p. 348, 1967.
31. Bland R. D., McMillan D. D., Bressac M. A., et al.:
Clearance of liquid from lungs of newborn rabbits. J. Appl.
Physiol. 49:171, 1980.
32. Bland R. D.: Dynamics of pulmonary water before and
soon after birth. Water
and electrolyte balance in newborn infants. Symposium, Departments
of Paediatrics and Paediatric Surgery, University of Uppsala, Sweden,
November 1314, 1980.
33. Civin W. W., Edwards J. E.: Postnatal structural
changes in the intrapulmonary arteries and arterioles. A. M. A.
Arch. Pathol. 51:192, 1951.
34. Naeye R. L.: Arterial changes during the perinatal
period. Arch. Pathol. 71:121, 1961.
35. Cook C. D., Drinker P. A., Jacobson H. L., et al.:
Control of pulmonary blood flow in the foetal and newly born lamb. J.
Physiol. (Lond) 169:10, 1963.
36. Lock J. E., Olley P. M., Coceani F., et al.: Use of
prostacyclin in persistent fetal circulation. Lancet 1:1343,
1979.
37. Wallgren G.: Den centrala cirkulationshostallningen vid
fodelsen. Lakartidningen 74:1708, 1977.
38. Kennedy J. A., Clark S. L.: Observations on the ductus
arteriosus of the guinea pig in relation to its method of closure.
Anat. Bee. 79:349, 1941.
39. Kennedy, J. A., Clark S. L.: Observations on the
physiological reactions of the ductus arteriosus. Am. J.
Physiol. 136:140, 1942.
40. Coceani F., Olley P. M.: The response of the ductus
arteriosus to prostaglandins. Can. J. Physiol. Pharmacol.
51:220, 1973.
41. Starling M. D., Elliott R. B.: The effect of
prostaglandins, prostaglandin inhibitors and oxygen on the closure of
the ductus arteriosus pulmonary arteries and umbilical vessels in
vitro. Prostaglandins 8:187, 1974.
42. Sharpe G. L., Larsson K. S.: Studies on closure of the
ductus arteriosus. In vivo effect of prostaglandin. Prostaglandins
9:703, 1975.
43. Sharpe G. L., Larsson K. S., Thalme B.: Studies on
closure of the ductus arteriosus. XII. In utero effect of
indomethacin and sodium salicylate in rats and rabbits.
Prostaglandins 9:585, 1975.
44. Lagercrantz H., Bistoletti P., Nylund L.:
Sympathoadrenal activity in the foetus during delivery and birth. In
Stern L. (ed.): Intensive Care in the Newborn. III. Baltimore:
Waverly Press, 1981.
45. Friedman W. F.: The intrinsic physiological properties
of the developing heart. In Friedman W. F., Lesch M., Sonnenblick, E.
H. (eds.): Neonatal Heart Disease New York: Grime &
Stratton, 1973, p. 21.
46. Comline R. S., Silver M.: Development of activity in
the adrenal medulla of the foetus and newborn animal. Br. Med.
Bull. 22:16, 1966.
47. Hervonen A., Korkola O.: The effects of hypoxia on the
catecholamine content of human fetal abdominal paraganglia and
adrenal medulla. Acta. Obstet. Gynecol. Scand. 51:17, 1972.
48. Lagercrantz H., Bistoletti P.: Catecholamine release in
the newborn infant at birth. Pediatr. Res. 11:889, 1977.
49. Jones C. T., Ritchie J. W. K.: The metabolic and
endocrine effects of circulating catecholamines in fetal sheep. J.
Physiol. (Lond) 285:395, 1978.
50. Eliot R. J., Lam R., Artal R., et al.: Norepinephrine
levels at birth and during the first 48 hours of life in the
human. Pediatr. Res. 12:412, 1978.
51. Dawes G. S.: Foetal and neonatal physiology. A
comparative study of the changes at birth. Chicago: Year Book
Medical Publishers, 1968.
52. Downing S. E., Gardner T. H., Rocamora J. M.: Adrenergic
support of cardiac function during hypoxia in the newborn lamb.
Am. J. Physiol. 217:728, 1969.
53. McPherson R. A, Kramer M. F., Covell J. W., et al.: A
comparison of the active stiffness of fetal and adult cardiac muscle.
Pediatr. Res. 10:660, 1976.
54. Riemenschneider T., Hirschfeld S., Riggs T., et al.:
Echographic ventricular systolic time intervals in normal term and
preterm neonates. Pediatrics 62:317, 1978.
55. Arcilla R. A., Lind J., Zetterqvist P., et al.: Hemodynamic
features of extrasystoles in newborn and older infants. Am. J.
Cardiol. 18:191,1966.
56. Hedvall G.: Systolic time intervals in newborn infants.
Acta. Pediatr. Scand. 64:839, 1975.
57. Lundell B. P. W., Wallgren G.: Left ventricular adaptation to
extrauterine circulation. Systolic time intervals in the newborn
infant. Acta Paediatr. Scand. (In press, 1981)
58. Cornblath M., Schwartz R.: Carbohydrate homeostasis in the
neonate. In Disorders of Carbohydrate Metabolism in Infancy. Series
in: Major Problems in Clinical Pediatrics. Philadelphia: W. B.
Saunders Company Vol. 3, 2nd ed. p. 72-87,1966.
59. Shelley H. J.: Glycogen reserves and their changes at birth.
Br. Med. Bull. 17:127,1961.
60. Stafford A., Weatherall J.: The survival of young rats in
nitrogen. J. Physiol. 153:457, 1960.
61. Oh W., Arcilla R. A., Lind J., et al.: Arterial blood gas and
acid base balance in the newborn infant: Effect of cord clamping at
birth. Acta Paediatr. Scand. 55:593, 1966.
62. Usher R., Shephard M., Lind J.: The blood volume of the
newborn infant and placental transfusion. Acta Paediatr. Scand.
52:497, 1963.
63. Oh W., Oh M. A., Lind J.: Renal function and blood volume in
newborn infant related to placenta transfusion. Acta Paediatr.
Scand. 56:197, 1966.
64. Oh W., Lind J., Gessner 1. H.: Circulatory and respiratory
adaptation to early and late cord clamping in newborn infants.
Acta Paediatr. Scand. 55:17, 1966.
65. Arcilla R. A., Oh W., Wallgren G., et al.: Quantitative
studies of the human neonatal circulation. II. Hemodynamic findings
in early and late clamping of the umbilical cord. Acta Paediatr.
Scand. Suppl. 179:25, 1967.
66. Yao A. C., Lind J., Vuorenkoski V.: Expiratory grunting in the
late clamped normal neonate. Pediatrics 48:865, 1971.
67. Yao A. C., Lind J.: Effect of early and late clamping on the
systolic time intervals of the newborn infant. Acta Paediatr.
Scand. 66:489, 1977.
Return to the Table of Contents Page
Created 9/14/2002 / Last modified 9/14/2002
Neonatology on the Web / webmaster@neonatology.net